Microbiology Daily Newsletter
Microbiology Daily Newsletter
January 28, 2014 - Metabolic Diversity
Prokaryotes have amazing metabolic diversity, unlike eukaryotes which are metabolically very similar. Bacteria and Archaea can utilize an incredibly diverse pool of carbon, nitrogen, phosphorus and sulfur compounds. Individuals have been shown to use unusual metals, produce compounds never thought to exist naturally, and even degrade compounds we believed were refractory (or non-biodegradable).
For example, 2,4,6-Trinitrotoluene (TNT) can be degraded by Pseudomonas spp. Below is a diagram showing proposed and investigated degradation pathways.
 |
| Esteve-Núñez A et al. Microbiol. Mol. Biol. Rev. 2001;65:335-352 |
From here, think about what you know of metabolism and organic chemistry (you thought that would never come up again...didn't you?). Follow the path leading down from TNA. Notice that like with Glycolysis and TCA, we are doing a step-wise alteration of the molecule, and we end with toluene. Toluene can enter the TCA cycle.
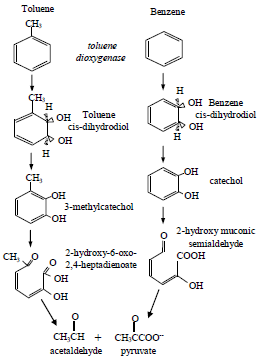 |
TRIGUEROS, D.E.G.; MODENES, A.N. y RAVAGNANI, M.A.S.S.. Biodegradation kinetics of benzene and toluene as single and mixed substrate: estimation of biokinetics parameters by applying particle swarm optimization. Lat. Am. appl. res. [online]. 2010, vol.40, n.3, pp. 219-226. ISSN 0327-0793. |
At this stage of your academic career, you should begin getting use to pathway diagrams; not only following them but understanding what is happening. Microbes, bacteria, archaea, and fungi all have unique and interesting metabolic pathways, and to understand them fully, you must be able to understand their metabolism.
Daily Challenge
Why you studied glycolysis, you studied the most common form, known as the Embden–Meyerhof–Parnas (EMP) pathway. The Entner-Doudoroff pathway is a variation of the standard glycolytic pathway that can be found in bacteria genera such as Pseudomonas, Escherichia, and Enterococcus. The lactic acid bacteria (a group, or more specifically, a clade of Gram positive bacteria) make use of a pentose-phosphate pathway during catabolism.Compare and contrast the Entner-Doudoroff and Pentose-Phosphate pathway with the typical EMP pathway you have previously learned. What are the advantages of these other catabolic pathways, and what are the limitations? What are important reactions, and why would certain organisms preferentially use these?