 Special Edition
Special Edition
October 25, 2012 DNA Replication
Textbooks have a tendency to make
replication one of the most complext topics covered. With a tendency to throw all the current research and understanding at students, they rarely take a step back and try to explain it. This newsletters has two goals: 1) to help biology students understand DNA replication, and 2) to show you want is expected from a Biology Freshman/Sophmore.
 |
| Central Dogma of Biology |
Before we get into replication, let's take a step back and look at the three core genetic processes, a.k.a., the Central Dogma. The central dogma describes the flow of
genetic information in a cell. The core idea is
INFORMATION. You may recall some of our
early discussion on DNA, and about base complimentarity and the directionality of the molecule. This will become important rather quickly, but for just this moment, I want you to concentrate on the fact
that DNA carries information. Information on how to build RNA and Proteins, both of which will produce the phenotype (expression) of the cell. For this reason, DNA, RNA and Proteins are considered
Informational Macromolecules. This means that the
sequence of monomers contains infomation,
e.g., instructions on how to build RNA and Proteins. Since it is critical, the
sequence of nucleotides carries information.
As you can see from the diagram of central metabolism shown above, there are three processes: Replication, Transcription and Translation. Think about those words. They are words used in reference to languages and documents. When you replicate a docuement, you want to ensure that you are getting a faithful (or even exact) copy of the original.
When you transcribe, you are moving from one medium,
e.g., spoken word, to another medium,
e.g., text. If you watch news shows, they will tell you that
transcripts of the show are available. Court reports make
transcripts of the trial. You are taking the language from one medium (in our case DNA) to another medium (RNA). The language is still the same (
i.e., nucleic acid lanugae), just in a different form. Does the transcript have to be 100% correct? You want it to be, but it is not as exacting as a replication.
Translation is where you change languages. Unless you're fluent in another language, you will need somoene to help translate, or at least a good translation dictionary. Now you are moving the context from one language (nucleic acid) to another language (amino acids).
All of these processes rely on one key feature of nucleic acids:
BASE COMPLIMENTARITY. In
DNA: A complements T, and G complements C. In
RNA: U complements A, and G still complements C.
Replication
In replication our
goal is to take
one molecule of DNA and make two daughter molecules of DNA that are identical to the first. Even the best replication processes can produce errors, but our goal is to be error free. This takes precision! As DNA is long, we also need this to be a fast process.
The enzyme that is ultimately responsible for replication is
DNA Polymerase.
[NOTE: there are multiple types of DNA Polymerase, but for now you just need to understand the core concept common to the DNA Polymerase family.] DNA polymerase is only one component of the
Replication Complex, which is a complex association of proteins needed to successfully complete the replication event. Your goal at this time is to concentrate on DNA polymerase; we will talk about some of the other components later.
. Available at: http://upload.wikimedia.org/wikipedia/commons/thumb/e/e4/DNA_chemical_structure.svg/206px-DNA_chemical_structure.svg.png. Accessed October 25, 2012.")
DNA is a double stranded molecule, in which the strands are anti-parallel. This means that one strand starts at the 5' end
→ 3' end, while the the other strand is revered. This can be seen in the image to the right. A common way of saying this is that we
read DNA in the 5'
→ 3' direction. Why is this important?
DNA Polymerase can only read DNA in the 5' → 3' direction, and can only build a new strand in the 3' → 5' direction. Before we go on, let's look another time at how deoxyribonucleotides are polymerized.

To the left you will see a generic image showing DNA Polymerase adding nucleotides to a growing DNA strand. Look carefully: A deoxyribonucleotide triphosphate (dTTP) is being added. The 5' phosphate of the new nucleotide is what will be used to form the phosphodiester bond. DNA Polymerase
requires a free 3' end on which to grow the new DNA strand. To the right is another image that will help you with this concept.
. Available at: http://www.bio.miami.edu/dana/pix/phosphodiester.jpg. Accessed October 25, 2012.")
At this point, the critical thing to remember is that DNA Polymerase will need a free 3' end on which to add a new nucleotide.
This requirement to build in only one direction (
3' → 5') creates a problem for the DNA process: the two strands read in opposite directions, and each must be replicated. It was noticed that one strand appears to replicate
continuously, while the opposite strand appears to replicate
discontinuously.
The original strand that reads 5'
→ 3' can be used by DNA polymerase to continuously produce the new 3'
→ 5' strand (the antiparallel complement to the original strand). We refer to the continuous synthesis strand as the
Leading Strand. The other original strand, which read 3'
→ 5' cannot be copied continuously. A section will have to be exposed, replicated, and then another section exposed. This strand is constructed discontinuously, and is reffered to as the
Lagging Strand.
 - Scaled (0%). Available at: http://25.media.tumblr.com/tumblr_mbs7jp80441qc0hbao1_1280.jpg. Accessed October 25, 2012.")
The image above shows the leading and lagging strand. Notice that the leading strand is replicating toward the
Replication Fork(where the original strands seperate). As more DNA unwinds and opens, DNA synthesis continues down the leading strand.
The lagging strand though has to build in a start-stop action, producing
Okazaki fragments. These fragments have to be sealed (phosphodiester bonds) together before DNA can rewind into the α-helix.
In the image above, you will notice a number of enzymes on the lagging strand. These enzymes are required for the initiation of DNA polymerazation, and then sealing the fragments. Looking at the image, you will see an enzyme called
Primase, and a structure in red known as a
Primer. Another restriction on DNA polymerase is that it must have a free 3' end from which to start building. DNA Polymerase is prevented from building a DNA strand from nothing. Something (a primer) must be in place upon which DNA polymerase can build. The Primer is constructed from RNA, and is a temporary scaffold upon which DNA polymerase can start working.
Eventually the primer will need to be moved. This is where you need to learn a little more about DNA polymerase. There are multiple DNA polymerases in eukaryotic systems. The general work horse of replication is
DNA Polymerase III (DNA pol III), which is used to make long strands of DNA.
DNA Polymerase I (DNA pol I) is used to replace primers (it is also used in repair functions). Even after the RNA primer is replaced with DNA, there is still a gap between fragments.
Ligase is the enzyme used to create a phosphodiester bond between fragments, thus sealing the new sugar-phosphate backbone of the synthesized strand.
The result of DNA replication is that one molecule one DNA was used to create two new molecules of DNA. The two strands of the original DNA molecule became the template from which to build new
complimentary strands of DNA. This is referred to as semi-conservative replication, as each new molecule has one strand from the original molecule, and one freshly synthesized
complimentary strand.
Optional Challenge
In your own words describe the purpose and process of replication. While you are encouraged to deepen your knowledge of replication, you are only required at this time to go into as much detail as was presented in this newsletter.
Link to Forum
Link to Lecture Reflection Forum


. Available at: https://wikispaces.psu.edu/download/attachments/46924776/image-2.jpg. Accessed October 30, 2012.") In eukaryotic DNA every gene starts with a promoter. This is a sight of ~8 nucleotides visible in the major groove of DNA. The transcription complex recognizes this sequence as a "START" indicator. The main core of the transcription complex will be RNA polymerase. This enzyme works to build a strand of RNA complementary to DNA.
In eukaryotic DNA every gene starts with a promoter. This is a sight of ~8 nucleotides visible in the major groove of DNA. The transcription complex recognizes this sequence as a "START" indicator. The main core of the transcription complex will be RNA polymerase. This enzyme works to build a strand of RNA complementary to DNA. 






 molecule. As can be seen in the image to the right, the major groove is wide enough to "see" the base pairs. The base pairs have an electrochemical profile, and thus can respond to other chemicals (via van Der Waals forces). Thus,
molecule. As can be seen in the image to the right, the major groove is wide enough to "see" the base pairs. The base pairs have an electrochemical profile, and thus can respond to other chemicals (via van Der Waals forces). Thus,  - Scaled (0%). Available at: http://deliciouslygrey.files.wordpress.com/2011/10/pdbimportvisualization2.png?w=300&h=300. Accessed October 29, 2012.") The image to the right shows a bacterial promoter event. One of the factors needed to start transcription (by recognizing the promoter) has bound into the major groove. This recognition event is needed to identify the start point of a gene. The Transcription Initation Complex will then begin to form at this site, and begin the transcription of the gene.
The image to the right shows a bacterial promoter event. One of the factors needed to start transcription (by recognizing the promoter) has bound into the major groove. This recognition event is needed to identify the start point of a gene. The Transcription Initation Complex will then begin to form at this site, and begin the transcription of the gene. . Available at: http://universe-review.ca/I11-37-operon.jpg. Accessed October 29, 2012.") House keeping genes are those that are needed for the general function of the cell, and can include genes for glycolysis, citric acid cycle, and ribosomes. These genes are always ON, and are referred to as
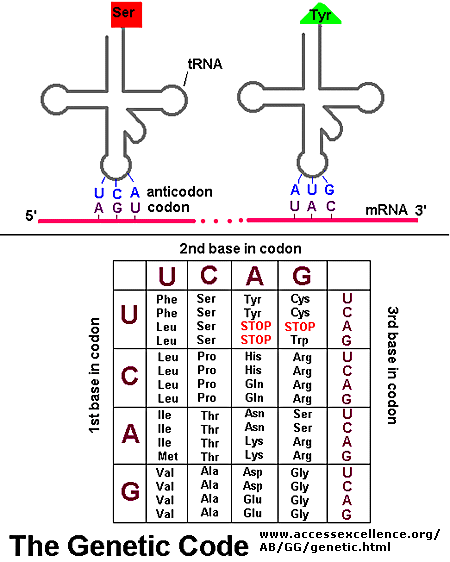
House keeping genes are those that are needed for the general function of the cell, and can include genes for glycolysis, citric acid cycle, and ribosomes. These genes are always ON, and are referred to as . Available at: http://2.bp.blogspot.com/-i-CYqAFf61o/TZXYoBQmXrI/AAAAAAAAB5g/5PmMdfOP1PQ/s1600/genetic-code-1.jpg. Accessed October 29, 2012.")
. Available at: http://bio3400.nicerweb.com/Locked/media/ch13/13_07-genetic_code.jpg. Accessed October 29, 2012.")
 - Scaled (20%). Available at: http://upload.wikimedia.org/wikipedia/commons/thumb/d/d6/GeneticCode21-version-2.svg/2000px-GeneticCode21-version-2.svg.png. Accessed October 29, 2012.")

. homolg.gif, found at http://staff.jccc.net/pdecell/celldivision/chromoterm.html, accessed on October 25, 2012.") one from their mother and one from their father. These two individual examples of chromosome 1 are considered homologous. Homologous chromosomes carry the same genes, but each homolog (one copy) will have a unique set of alleles (one allele for each gene).
one from their mother and one from their father. These two individual examples of chromosome 1 are considered homologous. Homologous chromosomes carry the same genes, but each homolog (one copy) will have a unique set of alleles (one allele for each gene).  - Scaled (0%). Available at: http://www.hartnell.edu/tutorials/biology/images/homologous_chromosomes.jpg. Accessed October 25, 2012.") IMPORTANT NOTE: Remember that what we are seeing are chomosomes. The two chromatids of a given chromosome are the products of DNA replication. The image to the left should help you remember that you start with a Maternal DNA molecule and a Paternal DNA molecule. After DNA replication, these will consense into maternal and paternal chrmosomes. It is critical that you remember the difference between Chromosome and Chromatid.
IMPORTANT NOTE: Remember that what we are seeing are chomosomes. The two chromatids of a given chromosome are the products of DNA replication. The image to the left should help you remember that you start with a Maternal DNA molecule and a Paternal DNA molecule. After DNA replication, these will consense into maternal and paternal chrmosomes. It is critical that you remember the difference between Chromosome and Chromatid.. Available at: http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Chromosomal_Recombination.svg/200px-Chromosomal_Recombination.svg.png. Accessed October 25, 2012.") Prophase 1 (prophase of Meiosis I) is the most critical stage of meiosis. Prior to the reduction division (which occurs in Anaphase 1), a
Prophase 1 (prophase of Meiosis I) is the most critical stage of meiosis. Prior to the reduction division (which occurs in Anaphase 1), a . Available at: http://users.path.ox.ac.uk/~pcook/students/Figs/803w.png. Accessed October 25, 2012.") This recombination event begins when homologous chromosomes are brought together during prophase/prometaphase I. The homologs possess DNA sequence similarities, and are able to bind to each other (the homologs chemically recognize each other). Regions where Cross-Over (recombination) can occur begin to over lap, forming a visible structure known as a
This recombination event begins when homologous chromosomes are brought together during prophase/prometaphase I. The homologs possess DNA sequence similarities, and are able to bind to each other (the homologs chemically recognize each other). Regions where Cross-Over (recombination) can occur begin to over lap, forming a visible structure known as a . SparkNote on Mitosis. Retrieved October 25, 2012, from http://www.sparknotes.com/biology/cellreproduction/mitosis/")

 NOTICE: each cell produced in meiosis I now undergo meiosis II.
NOTICE: each cell produced in meiosis I now undergo meiosis II.  Daily Newsletter
Daily Newsletter. Cell Cycle 2-2.svg. Found at http://en.wikipedia.org/w/index.php?title=File:Cell_Cycle_2-2.svg&page=1, accessed on October 24, 2012.") Interphase is divided into three distinct steps: G1 Phase, S Phase, and G2 Phase.
Interphase is divided into three distinct steps: G1 Phase, S Phase, and G2 Phase.
. gem1s4_9.jpg. Found athttp://www.mhhe.com/biosci/ap/ap_prep/bioD4.html, accessed on October 24, 2012.") Throughout the cell cycle, there are time points referred to as
Throughout the cell cycle, there are time points referred to as 


. SparkNote on Mitosis. Retrieved October 23, 2012, from http://www.sparknotes.com/biology/cellreproduction/mitosis/") Once the chromsomes have been arranged, they can be seperated.
Once the chromsomes have been arranged, they can be seperated. . SparkNote on Mitosis. Retrieved October 23, 2012, from http://www.sparknotes.com/biology/cellreproduction/mitosis/")
. SparkNote on Mitosis. Retrieved October 23, 2012, from http://www.sparknotes.com/biology/cellreproduction/mitosis/")
 - Scaled (0%). Available at: http://images.tutorvista.com/content/feed/tvcs/cytodraw.gif. Accessed October 23, 2012.") Plants and fungi, which posses cell walls, experience cytokinesis in a dramatically different way. Prior to division, the plant cell will have made numerous vesicles that contain the raw materials needed to create cell walls. These materials are held in an inert state, with inactive enzymes needed to form these materials into walls. As telophase begins, these vesicles begin to line up down the equator of the cell, and begin to fuse (what do you think triggers this action? will it trigger the enzymes?). As the vesicles fuse, cell walls begin to form. As more vesicles fuse (remember the membrane is dynamic), the wall continues to grow (
Plants and fungi, which posses cell walls, experience cytokinesis in a dramatically different way. Prior to division, the plant cell will have made numerous vesicles that contain the raw materials needed to create cell walls. These materials are held in an inert state, with inactive enzymes needed to form these materials into walls. As telophase begins, these vesicles begin to line up down the equator of the cell, and begin to fuse (what do you think triggers this action? will it trigger the enzymes?). As the vesicles fuse, cell walls begin to form. As more vesicles fuse (remember the membrane is dynamic), the wall continues to grow (. Apoplast and symplast pathways.svg. Found at http://en.wikipedia.org/w/index.php?title=File:Apoplast_and_symplast_pathways.svg&page=1, accessed on October, 23, 2012.") NOTE: Plant cell walls are generally impervious to water flow. As a by-product of plant cytokinesis, and the development of the cell plate, you will find holes between the two new daughter cells. Known as plasmodesmata, these membrane lined holes allow for a continuous cytoplasm between cells. This allows for rapid movement of water and nutrients (including plant hormones) between cells (
NOTE: Plant cell walls are generally impervious to water flow. As a by-product of plant cytokinesis, and the development of the cell plate, you will find holes between the two new daughter cells. Known as plasmodesmata, these membrane lined holes allow for a continuous cytoplasm between cells. This allows for rapid movement of water and nutrients (including plant hormones) between cells (


